クロアシオキノタユウ(クロアシアホウドリ)

写真:長谷川 博
種名
| 和名 | クロアシオキノタユウ(クロアシアホウドリ) |
|---|---|
| 学名 | Phoebastria nigripes |
| 英名 | Black-footed Albatross |
絶滅危険度
日本(環境省):指定なし 世界(IUCN):準絶滅危惧種(NT) |
法的保護
主な繁殖地である伊豆諸島鳥島は文化財保護法の「天然保護区域」として天然記念物に指定され(1965年)、この島に文化庁からの許可なく上陸することは禁止されている。また、鳥島は環境省によって国指定鳥獣保護区(特別保護地区)に指定されている。この種は小笠原諸島でも繁殖し、繁殖地となっている島嶼は「国指定小笠原群島鳥獣保護区(特別保護地区)」に含まれ、また、国立公園の特別保護地区に指定(1972年)されている。さらに、世界自然遺産にも登録された(2011年)。琉球諸島の尖閣諸島南小島・北小島にも繁殖地があるが、これらの島嶼への立ち入りは国によって禁止されている。
個体数減少の原因
かつて伊豆諸島鳥島や小笠原諸島聟島列島、尖閣諸島などで繁殖していたが、オキノタユウ類としてまとめて(種ごとに区別しないで)記述されることが多かったため、この種の個体数がどれほどであったか、はっきりしない。ただ、鳥島では海岸に近い斜面で繁殖していて、島の中腹以上で密集した大集団をなして繁殖していたオキノタユウと較べると、個体数ははるかに少なかったにちがいない。おそらく、その他の島嶼でも同様だっただろう。
この種も1880年代末から始まった羽毛採取のために捕獲されが、その羽毛は黒褐色から灰褐色で、オキノタユウの白い羽毛よりも低価であったため、激しい捕獲を免れた。鳥島では1932年に700〜800羽が観察され、1933年に鳥島は禁猟区(10年間)に指定されたが、監視の目が行き届かない離島で捕獲が続き、1939年には約30羽にまで減少した。その夏に鳥島の火山が大噴火を起こし、大量の火山灰と溶岩を噴き出し、営巣地の環境を変えた。その結果、繁殖活動が影響を受けたにちがいない。小笠原諸島の聟島列島も1936年に禁猟区(10年間)に指定されたが、その効果は定かではない。尖閣諸島では1900年代初頭に久場島や南小島・北小島で繁殖していたが、捕獲によって個体数が減少した。戦後、1960年代まで、台湾の漁民がこれらの島に上陸し、海鳥類の卵を採取した。そのとき成鳥やひなも捕獲されたと思われる。
日本列島の繁殖地では個体数が増加してきたが、この種のおもな繁殖地である北西ハワイ諸島(約60,000組のつがいが繁殖)では、それまで増加傾向にあった個体数が1990年代から減少傾向に転じ、その主な原因は漁業、とくに延縄漁による混獲であると判明した。そのため1998年に、国際鳥類保護団体バードライフ・インターナショナルはこの種を危急種(VU)とし、さらに2008年の絶滅危険度評価では1ランク上げて絶滅危惧種(EN)とし、2014年にもそれを維持した。しかし21世紀になって、延縄漁による混獲を防止・軽減する手法(たとえば「鳥ライン」)が確立し、それらが操業の現場で広く取り入れられ、漁業による混獲が減少して個体数回復の傾向がみえたため、2019年には2ランク下がって準絶滅危惧種(NT)になった。
北西ハワイ諸島では、海上に浮遊するプラスチックごみ(その小片)を成鳥が大量に「誤食」して消化管を詰まらせて死亡する例が観察される。また、ひなも親鳥からプラスチック類が混入したえさを与えられて死亡する例が報告されている。今後、プラスチック類による海洋汚染がさらに深刻化すれば、個体数減少の原因の一つになるだろう。
日本での保護活動の歴史
伊豆諸島鳥島では1950年代半ばから気象観測所の職員によって、オキノタユウの繁殖状況監視調査や保護活動と並行して、この種の繁殖状況も調査された。この期間に、ひなを捕食するおそれのある野生化したネコの駆除が行なわれたが、その他の特別な保護対策は講じられなかった。
1956-57年繁殖期には20羽近い成鳥が観察され、6羽のひなが巣立ち、1959-60年期には34羽が観察され、10羽のひなが巣立った。そして、1964-65年期には83羽が観察され、30組のつがいが産卵し、20羽のひなが巣立ち、順調に個体数が増加した。しかし、1965年の秋に気象観測所が閉鎖されて無人になり、監視調査と保護活動はとだえた。
1973年に、イギリス人鳥類学者が鳥島に上陸して調査し、50羽のひなを観察し、この繁殖集団の成長が確認された。その後、1976年期から85年期まで不定期に、1988年期からは毎年、ひなのセンサス(全数調査)が行なわれ、鳥島集団のきわめて順調な成長が明らかになった(図1。当てはめた曲線はおおまかな傾向を示す)。

図1.伊豆諸島鳥島におけるクロアシオキノタユウ集団の成長(まとめ:長谷川博)
かつて、小笠原諸島聟島列島にノヤギが持ち込まれ(食用目的)、野生化して数多く生息していた。その結果、生息している島々(聟島、媒島、嫁島)の植生が破壊され、土壌の浸食が起こった。この種はノヤギによる撹乱(営巣妨害)をさけて、ノヤギのいない島嶼で繁殖していた。1997年からノヤギの排除が始められ、2006年に完了した。その結果、この種はノヤギが排除された島で繁殖するようになり、近年、個体数が増加し、営巣分布域が拡大している。
沖縄の尖閣諸島では、1939年に北小島でたくさんいたと記録されているが、戦後は生息が確認されなかった。ようやく1970年に北小島で少数が観察され、1971年に南小島と北小島で数羽が確認された。2002年の調査では、北小島の上部の岩場で見える範囲にひな37羽、成鳥・若鳥89羽、南小島の頂上部の斜面でひな5羽、成鳥・若鳥36羽が観察された。両島を合計するとひなは少なくとも42羽、成鳥・若鳥は少なくとも125羽となり、個体数の増加が確認された。その後、国が尖閣諸島への立ち入りを禁止したため、繁殖状況の調査は困難になった。
繁殖分布と個体数の現状と動向
伊豆諸島鳥島では、1950年代から1980年代半ば過ぎまで、この種は島の南東端に位置する燕崎の斜面で、オキノタユウの営巣地より下のなだらかな砂礫地で営巣していた。ここでの繁殖つがい数が急速に増えた結果、1980年ころから島の北西側のなだらかな斜面にごく少数がたまに見られるようになった。そして、1987年の秋に火山灰が堆積した燕崎斜面で地滑り(表層崩壊)が起こり、泥流が営巣地の一部に流入したことがきっかけとなって、1988年の秋に4組のつがいが北西側の斜面で産卵し、翌春にそこから初めて1羽のひなが巣立った。
その後、繁殖つがい数が順調に増えて、1999年の秋には燕崎の崖上にある平地、北西側斜面の兵庫浦上方でも繁殖し始めた。さらに2005年からは気象観測所跡の南上方、2008年からは兵庫湾の上方でも繁殖を開始し、島の西から北の斜面に営巣分布域を急速に拡大した。2018年の春、大きく成長したひなの数は、燕崎斜面に530羽、その崖上地区に84羽、北西斜面に1,617羽、兵庫浦上方地区に360羽、気象観測所跡の南上方地区に179羽、兵庫湾上方地区に100羽、合計2,870羽であった。このひな数から繁殖つがい数をおおまかに推定すると、当時の鳥島集団はおよそ4,500組、総個体数は約25,000羽となる。この集団の成長が鈍化する明らかな兆候はまだなく(図2)、今後しばらくの間、このまま成長するだろう(年増加率は6.6%)。
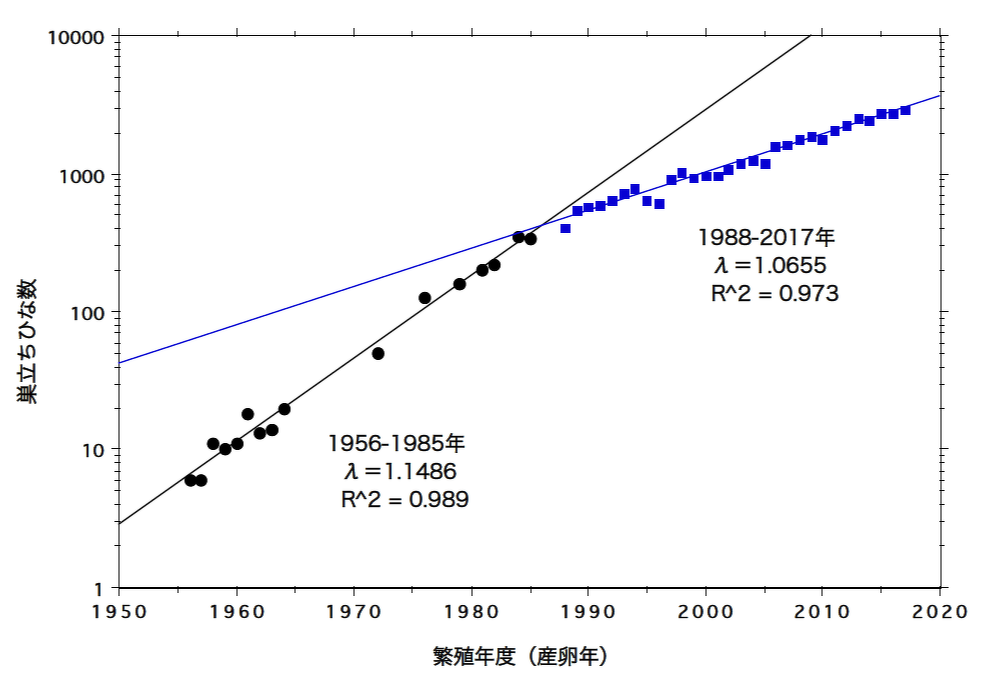
図2. 伊豆諸島鳥島におけるクロアシオキノタユウ集団の成長率:北西側斜面への新営巣地形成(1988年)の前後で比較
伊豆諸島の八丈小島では、以前にノヤギが生息していたが、植生の破壊が進んで土壌の浸食が起こり、沿岸海域の生物群集や漁業にも影響が現れたため、2001年からノヤギの排除が行なわれ、2007年に完了した。その結果、2010年代になって、島の北部の平坦な区域でこの種が目撃されようになり、2013年に初めて産卵した。その後、繁殖集団は着実に増加し、数十組のつがいが繁殖するようになった。
小笠原諸島聟島列島では、1977年の聟島鳥島におけるコオキノタユウの繁殖の発見がきっかけになり、聟島鳥島および主要3島(聟島、媒島、嫁島)で東京都小笠原支庁による標識調査が開始され、2004年以降は(特非)小笠原自然文化研究所との共同事業として継続されている。1992年の調査で、クロアシオキノタユウは聟島列島の北之島、聟島、聟島鳥島、媒島、嫁島で繁殖していることが観察され、2001年の春には合わせて330羽以上のひなが確認された。上記の主要3島では、ノヤギによる植生の破壊が進行していたため、1997年からノヤギの排除が行なわれ、2001年に完了した。
ノヤギ排除の最終段階にあった聟島で、2001 年に確認されたひな数はわずか1 羽だったが、2016 年には6 羽になった。媒島では、2002 年にひな数が 117 羽だったが、ノヤギ排除から16年後の2017年には512羽にまで増加した。また、嫁島では2001 年にノヤギ排除が完了し、ひな数は78羽だったが、それから11 年後の 2012 年には 330 羽に、さらに2017 年には 413 羽にまで増加した。2017年時点で、これら主要3島に加えて、周辺の属島および北之島を含めた聟島列島の合計ひな数は、約1,200羽になった。その後、ひな数の増加傾向はほぼ止まり、2020年にはやや減少した。
聟島列島での繁殖集団の急速な成長にともなって、クロアシオキノタユウは繁殖分布域を拡大し、他の列島のいくつかの属島で繁殖を開始した。父島列島の孫島では2010年の春、母島列島の姉島南鳥島と妹島鳥島では2002年の春に新規定着が確認された。その後、孫島ではひな数がゆるやかに増加し、2018年に20羽になった。また、姉島南鳥島と妹島鳥島では、2018年に合わせて15羽のひなが確認された。その他の属島においても、飛来や営巣について数例の報告がある。
小笠原諸島におけるクロアシオキノタユウは、主要繁殖地の聟島列島で2001年にノヤギが排除されたことを機に、繁殖集団が急速に成長し、繁殖分布域を他の列島へと拡大してきた。しかし2017年以降、繁殖集団の成長は止まり、停滞状態にある。これが一時的現象なのか、それとも明らかな変化なのか、注意深く分析する必要がある。
尖閣諸島では繁殖状況の調査が行なわれていないので、くわしいことはわかっていない。ここでは上陸・立ち入りが禁止され、人間の影響をまったく受けていないので、繁殖集団は確実に成長しているはずである。
繁殖集団間の移動
1950年代から1980年代にかけて鳥島集団は平均して毎年、14.9%ずつ指数関数的に成長した(5年で倍加。図2参照)。このような急激な個体数の増加は他集団からの移入なくして実現不可能である。この期間にどの繁殖集団から移入したかは不明である(足環標識個体は確認されなかった)。
その後、小笠原諸島聟島列島で足環標識された複数の個体の鳥島集団への移入が確認された。最近も小笠原諸島で標識された個体が鳥島で観察されているので、小笠原諸島集団と鳥島集団との間で個体の交流が起こっていることは疑いない。
生態
大型の海鳥で、体重は約3kg、翼の開張は2.1mにも及ぶ。おもに大陸棚周縁部から外洋域を広範囲に移動して、イカ類や魚類、甲殻類を捕食し、浮遊している海洋動物の新鮮な死体や魚類の卵をも摂食する。伊豆諸島鳥島では、10月下旬に営巣地に帰還し、11月中旬から12月下旬に1卵を産む。雌雄交代で約65日間にわたって抱卵し、翌年1月半ば過ぎから2月中旬にひなが誕生する。ひなは両親に保育され、6月に巣立つ。巣立ち後、主に北太平洋の中緯度海域で非繁殖期を過ごす。
巣立ちから2、3年後に繁殖地に帰り、頻繁に求愛行動を行なってつがいを形成する。繁殖開始は早くて5歳(産卵から5年後)で、平均では7歳くらいだろう。毎年繁殖し、配偶システムは生涯一夫一妻で、死別するまでつがいの絆は維持される(死別後に再婚)。鳥島での繁殖成功率や巣立ち後の生残率は、まだ研究されていない。
個体数に影響を及ぼすおそれのある要因
- 漁業による混獲。この種が採食する海域で、1990年代から底延縄漁による混獲が増え、死亡率が高まって、北西ハワイ諸島の繁殖集団の個体数が減少した。この混獲事故を防止・軽減するため、海鳥類が延縄に近づかないようにする「鳥ライン」が開発され、この手法やその他の混獲防止措置が操業現場で実行されて、混獲による犠牲が大幅に減少した。今後も、採食海域での混獲防止措置が継続されれば、漁業による事故死は最小限に抑えられる。
- プラスチック類による海洋汚染。分解しにくいプラスチック類のごみは陸域から海洋に流れ込んで水面に浮遊し、大規模な海洋汚染を引き起こした。多くの海鳥類はそれらの小片をえさとまちがえて摂食し、大量に取り込んだ場合には消化管を詰まらせて死亡することがある。また少量であっても、プラスチック類の表面に吸着された化学物質が胃の中で溶け出し、体内に吸収されて健康被害を引き起こすことが明らかになっている。
- 火山噴火。伊豆諸島鳥島は日本列島でもっとも活発な火山(13山)の一つで、1902年と1939年に起こった大噴火は島の景観を変えた。もし、再び同規模の噴火が起これば、営巣地は大きな影響を受け、火山活動が終息するまで鳥島集団の繁殖活動は休止するだろう。しかし、火山噴火が及ぼす個体数への影響は一時的で、それによって継続的に個体数が減少することはない。
- 国際紛争。沖縄県の尖閣諸島に対して、中国と台湾は領有権を主張している。もし、領土をめぐる国際紛争が勃発すれば、南小島と北小島の営巣地にも影響が及ぶ。それを避けるために、政府間の対話による平和的解決が望まれる。
- 気候変動。地球規模での気候変動は、海洋環境を変質させ、海洋生態系を大きく変え、海鳥類の個体数に影響を及ぼすにちがいない。北西ハワイ諸島では、この種は海岸に近い低い場所で営巣するので、気候変動の影響で海水準が上昇すれば、発達した熱帯低気圧の高波によって営巣適地が失われるだろう。
主な保護課題
- 各地の繁殖集団を監視調査し、個体数の動向を把握する。上陸調査が不可能な営巣地をドローンで空中から撮影し、繁殖状況を把握する。
- 繁殖期・非繁殖期の採食海域を解明し、漁業による混獲を防止して海洋における保護を推進する。
- 繁殖成功率や巣立ち後の死亡率、繁殖開始年齢、繁殖参加率など、生活史の基本的特性を明らかにする。
執筆者
長谷川博(東邦大学名誉教授)
鈴木 創(小笠原自然文化研究所)
参考文献・資料
- Arata, J. A., Sievert, P. R. & Naughton, M. B. 2009. Status assessment of Laysan and Black-footed Albatrosses, North Pacific Ocean, 1923-2005. U. S. Geological Survey Scientific Investigations Report 2009-5131. 80pp.
- 鈴木創・堀越和夫・佐々木哲朗・川上和人. 2019. 小笠原諸島聟島列島におけるノヤギ排除後の海鳥営巣数の急激な増加. 日本鳥学会誌, 68: 273–287.